CONTENTS
INTRODUCTION
1. THE MAIN COMPONENTS OF THE BSB
2. COMPONENTS OF THE BSB MICROENVIRONMENT
CONCLUSIONS
COMPLIANCE WITH ETHICAL STANDARDS
FUNDING
CONFLICT OF INTEREST
Figure 1A comprehensive representation of the blood-salivary barrier.
Figure 2Intercellular contacts (tight junctions, gap junctions, adherents’ junctions and desmosomes).
Figure 3Pathologies associated with damage to intercellular junctions of the salivary glands and the oral cavity epithelium. Cx-43 – connexin 43, LSR – lipolysis-stimulated lipoprotein receptor, ZO-1 – zonula occludens, NF-κB – nuclear factor kappa-light-chain-enhancer of activated B cells, INF-y – interferon gamma, JAK – Janus kinases, STAT – signal transducer and activator of transcription, miR-145 – microRNA-145.
Figure 4Oral immune system. PRRs - pattern recognition receptors, PAMPs - pathogen-associated molecular patterns, DAMPs - damage-associated molecules, TNF-α - tumor necrosis factor-alpha, NF-κB - nuclear factor kappa-light-chain-enhancer of activated B cells, TLRs - Toll-like receptors, CLRs - C-type lectin receptors, NLRs - nucleotide oligomerization domain-like receptors, RLRs - retinoic acid-inducible gene I-like receptors, ALRs - AIM2 (Absent in melanoma 2)-like receptors.
Table 1Salivary gland SLC transporters
Table 2Intercellular junction proteins
|
THE BSB AS A MULTIFACETED SYSTEM: A REVIEW
E.I. Dyachenko, E.A. Sarf, L.V. Bel’skaya*
Key words: BSB; salivary glands; oral mucosa; microbiome; immune system; intercellular connections
Abbreviations: AJ – adherens junction; ALRs – AIM2-like receptors; aPKC – atypical protein kinase C; AQP5 – aquaporin 5; Clnd – claudin; CLRs – C-type lectin receptors; Cx – connexin; DAMPs – damage-associated molecules; ED – excretory duct; FNB6 – TERT2 immortalized cell line of oral keratinocytes derived from the buccal mucosa; HCO₃⁻ – bicarbonate; ID – intercalated duct; IFN-γ – interferon-gamma; IL – interleukin; JAK – janus kinase; JAM – junctional adhesion molecule; KRS – Krebs ringer bicarbonate solution; LSR – lipolysis-stimulated lipoprotein receptor; MALT – mucosa-associated lymphoid tissue; miR-145 – tumor-suppressing microRNA; Mist1 (Bhlha15) – a basic helix-loop-helix transcription factor found primarily in exocrine secretory cells such as pancreatic acinar cells, zymogenic cells of the stomach, and Paneth cells, regulates downstream genes that control se-cretory vesicle maintenance and trafficking; MUC – gene coding a family if proteins mucine; NF-κB – nuclear factor kappa-light-chain-enhancer of activated B cells; NLRs – nucleotide oligomerization domain-like receptors; NOD – non-obese diabetes; NOF – normal oral fibroblasts; NOK – normal oral keratinocytes; Ocln – occludin; PAMPs – pathogen-associated molecular patterns; PG – parotid salivary gland; PI3K – Phosphoinositide 3-kinase; Plg – plasminogen; Pm – plasmin; PRRs – pattern recognition receptors; RLRs – retinoic acid-inducible gene I-like receptors; SARS-CoV-2 – severe acute respiratory syndrome coronavirus 2; SD – striated duct; SLCs – solute carrier transporters; SLG – sublingual salivary gland; SMG – submandibular salivary gland; STAT – signal transducer and activator of transcription; TJ – tight junctions; TLRs – toll-like receptors; TNF-α – tumor necrosis factor-α; ZO – ZONULA occludens
The constancy of the body's internal environment is maintained by numerous physiological mechanisms, in which various histohematic barriers play an important role. Histohematic barriers include the blood-brain barrier, the blood-placental barrier, the blood-ocular barrier, and the blood-salivary barrier (BSB) [1-3]. The functional properties of the BSB are still require deeper studies and better understanding [4-6]. To date, a fully adequate model of the BSB, capable of accurately describing the causes and mechanisms of dynamic changes in its functional properties, has not been developed yet [7-10]. Furthermore, an interesting area of research is the study of the migration of molecules from blood to saliva and from saliva to blood depending on the integrity of the BSB, its physicochemical properties, and humoral and neural regulation in both physiological and pathological conditions [11-12]. Understanding the functional properties of the BSB in various pathologies, including cancer, will enable the use of saliva as an adequate and reliable biological material for diagnostics [13-15].
This work aims to provide a comprehensive analisys of the components associated with the BSB as well as known and completely new approaches used to study the BSB in vitro.
In this review, we consider the BSB as a complex system comprising both its core components and its microenvironmental components. The core components include the salivary glands and oral epithelium, the intercellular junction proteins responsible for barrier integrity and strength, saliva, and mucus. The microenvironmental components include the blood supply and innervation of the BSB, local immunity, and the oral microbiome. The microenvironment of the BSB directly influences its integrity and functional properties. In many ways, the structural components and the microenvironment are inseparable components of a single whole—the BSB (Fig. 1).
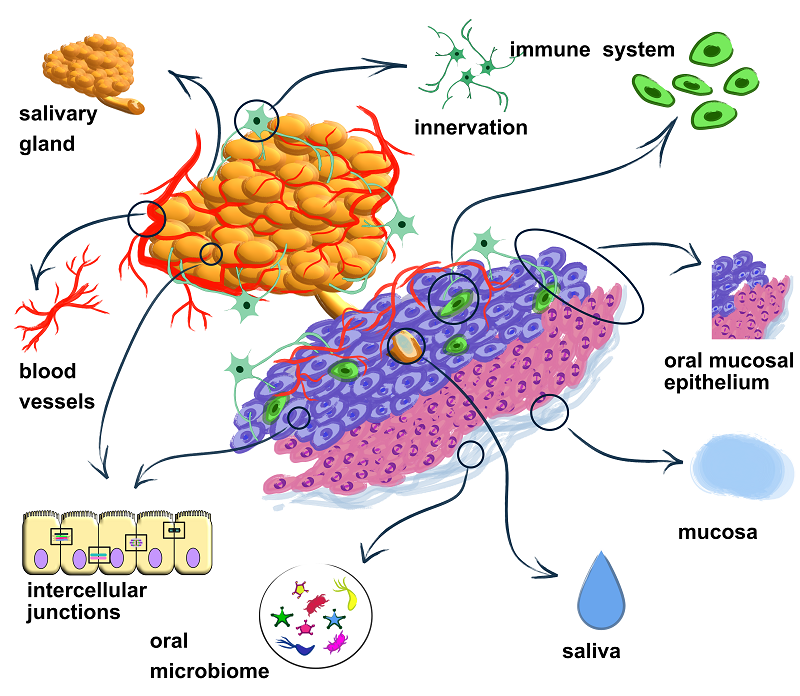
|
Figure 1.
A comprehensive representation of the blood-salivary barrier.
|
The oral mucosa epithelium is a biological barrier that serves as a boundary between the underlying tissues and the environment. In the context BSB, the oral mucosa epithelium acts as a barrier between the blood (internal environment) and saliva and the microbiome (external environment). The epithelium consists of a superficial stratified squamous epithelium and a dermis, which is represented by a loose papillary layer and a dense reticular layer [16]. The oral mucosa epithelium can be of two types: keratinizing and nonkeratinizing [17]. The oral mucosa epithelium performs a number of biologically important functions, such as: protective, plastic, sensory, and absorptive [18]. The BSB Permeability for drugs occurs mainly through the epithelium of the oral mucosa. Removal of lipids from the buccal mucosa leads to increased permeability for drugs [19]. There are significant differences in permeability between different areas of the oral cavity due to the different structures and functions of the different oral mucosa. In general, the permeability of the oral mucosa decreases in the following order: the sublingual mucosa is greater than the buccal mucosa, and the buccal mucosa is greater than the palatine mucosa [18].
The salivary glands are among the most complex and multifunctional components of the BSB. This is due to the fact that the salivary glands, like oral mucosa epithelium, perform separation functions between blood, saliva, and the microbiome, and also produce saliva and oral mucus [20-21]. Saliva and oral mucus themselves, in turn, perform barrier functions. There are the parotid, submandibular, sublingual, buccal, labial, and lingual glands in the oral cavity, and the parotid salivary gland (PG), the sublingual salivary gland (SLG), and the submandibular salivary gland (SMG) are three paired major salivary glands. The salivary glands have a branched structure with complex and numerous ducts. The terminal unit of the salivary glands is the acinus, which can be serous (producing more of the protein component), mucous (producing mainly mucins), or mixed serous-mucous. Serous acinar cells produce more protein; the cells are characterized by abundant eosinophilic zymogen granules and a spherical nucleus [22]. Mucous acinar cells have a transparent cytoplasm rich in mucins; the nuclei are polarized toward the cell basement membrane [23]. Acinar cells produce and secrete saliva, which is further modified and transported by a sequential duct system: intercalated duct (ID) cells, striated duct (SD) cells, and excretory duct (ED) cells, which connect the gland to the oral cavity [24]. The acini themselves are formed by 8–12 pyramidal acinar cells, which are laterally connected to each other by adhesive and tight junctions. Such lateral connections maintain the apical-basolateral polarity of the cells and prevent free lateral transport of ions between acinar cells. For ion transport, acinar cells contain specialized water channels and ion pumps, which regulate the secretion and concentration of salivary ions [25]. Identification of acinar cells occurs through the expression of markers such as aquaporin 5 (AQP5) and Mist1 (Bhlha15). Saliva is also secreted into the oral cavity by minor salivary glands located in the lips, cheeks, palate, tongue and respiratory tract [26-29].
Although the salivary glands have been well studied, a breakthrough was made in this area in 2020. Valstar et al. described the tubarial salivary glands [30], which are located between the nasal cavity and throat, deep in the nasopharynx, behind the tubal ridge (torus tubarius) [31-33]. Alisha Ebrahim et al. demonstrated that there are sex differences in the tubarial salivary glands, which explains gender differences in salivation. Thus, comprehensive criteria and approaches to the classification of salivary glands were revised to better describe histological features [34].
Within the BSB, such discoveries are important because they provide a more complete understanding of the properties of each component. This is useful for creating more adequate in vitro models, bioengineered oral mucosal equivalents, drug delivery, and the development of more gentle therapies for head and neck cancer and other diseases, all aimed at reducing alterations to the salivary glands and oral mucosa. For example, hypofunction of the salivary glands, which may be due to systemic disease (autoimmune diseases [35–36], diabetes mellitus [37–39], and thyroid diseases [40–41], neurological/psychiatric disorders [42–45], infectious [46–47], and genetic diseases [48–51], hypertension [52], and sarcoidosis [53]), medications [54], aging [55], and radiation therapy for head and neck cancer, can cause dry mouth, which increases the risk of diseases such as periodontitis, taste disturbances, pain and burning in the mouth, and dental caries [56]. A detailed description of the dysfunction of the salivary gland system in the above-mentioned diseases is the subject of a separate review; however, the general principles underlying these disorders include damage to the parenchyma of the salivary gland due to activation of the immune system and an increase in the level of reactive oxygen species, disruption of the innervation of the salivary glands, which directly affects its secretory activity, etc.
The principles of saliva secretion are well described in the literature. Furthermore, SLC transporters are known to play an important role in salivary gland secretion. These transporters are responsible for the transport of ions such as nitrites, chlorides, bicarbonates, and water. The functional activity of SLCs results in the formation of nitric oxide [57-58]. Currently known SLC transporters expressed by the salivary glands are presented in Table 1.
|
CLOSE

|
Table 1.
Salivary gland SLC transporters [ 57-60]
|
The most studied to date are SLC17A5, SLC4A2, SLC2A4, and SLC26A6 transporters. Involved in nitrate (NO₃⁻) entry SLC17A5 is located in the plasma membrane of salivary gland cells. It is important for the nitrate-nitrite-nitric oxide pathway, which facilitates nitric oxide signaling in mammals [59-60]. SLC4A2 is an electroneutral transporter. It is involved in the exchange of chlorides (Cl⁻) and bicarbonates (HCO₃⁻), thereby facilitating the regulation of intracellular pH and chloride concentration in salivary gland cells [60]. SLC2A4 and SLC26A6 are involved in the secretion of iodide (I⁻) and bicarbonate (HCO₃⁻); they also regulate the transmembrane conductance in cystic fibrosis in the parotid gland duct [60]. In the salivary glands SLC15A2, SLCO1A2, SLC22A1, SLC22A3, SLC22A4, SLC22A5, SLC22A6 and SLC22A7 transporter are expressed [60]. In the human salivary gland the SLC5A5 gene expression and the level of its protein product NIS (sodium/iodide symporter) are higher than that SLC17A5 NIS mediates the absorption of nitrate by the salivary glands and the concentration of nitrate in saliva [61]. Nitrate, through conversion to nitrite, forms nitric oxide, which is involved in signaling. Exposure to potassium iodide reduced the level of SLC5A5 in ductal salivary gland cancer cells, as well as their proliferation due to increased levels of reactive oxygen species and the triggering of caspase-dependent apoptosis [62].
The intercellular junction proteins, which include tight junctions, gap junctions, adherens junctions, and desmosomes are other important structural component of the BSB [63-65] (Fig. 2).
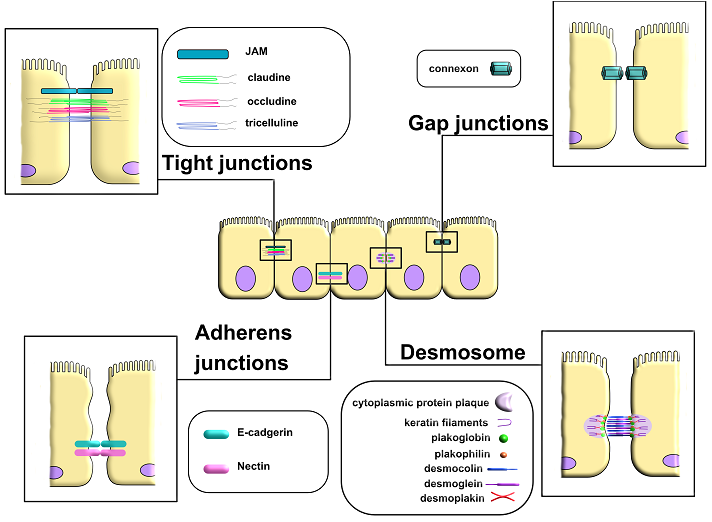
|
Figure 2.
Intercellular contacts (tight junctions, gap junctions, adherents’ junctions and desmosomes).
|
Many pathogens and somatic diseases lead to disruption of the structure and, consequently, dysfunction of tight and adherens junctions. This, in turn, further aggravates the condition of the area affected by the pathogenic change, leading to the progression of pathological processes [66]. A detailed description of the functions and structure of individual proteins of intercellular junctions is provided in Table 2.
|
CLOSE
|
Table 2.
Intercellular junction proteins
|
Disruption of contacts is observed in a number of pathologies. For example, in Sjogren's syndrome, the expression of epithelial tricellulin is suppressed by IFN-γ via the JAK/STAT1/miR-145 signaling pathway. It was shown that restoration of TJ integrity and re-expression of tricellulin resulted in normalization of salivation in patients [119]. Results of another study showed that increased IL-17 activity led to damage of tricellulin via the NF-κB signaling pathway in the submandibular salivary glands of mice with Sjogren's syndrome. In addition, the expression of claudin-1 and -3 increased, especially on the basolateral membranes, whereas claudin-4, occludin, and zonula occludens-1 (ZO-1) were decreased in the submandibular salivary glands of mice with non-obese diabetes (NOD) [120]. Certain evidence exist that the BSB was disrupted. In the disease this resulted in dilation of blood vessels and infiltration of lymphocytes, involved in the pathogenesis of Sjogren's syndrome [121]. The inflammatory environment resulted in disrupted localization and increased levels of claudin-5. Furthermore, a large accumulation of blood vessels was observed surrounding infiltrated lymphocytes [122-123]. Nishida et al. have demonstrated that lipolysis-stimulated lipoprotein receptor (LSR), localized in tight tricellular junctions and associated with lipid metabolism, also plays an important role in maintaining epithelial homeostasis. This receptor is highly expressed in normal cells and well-differentiated tumors and its expression decreases during tumor malignancy [124] (Fig. 3).
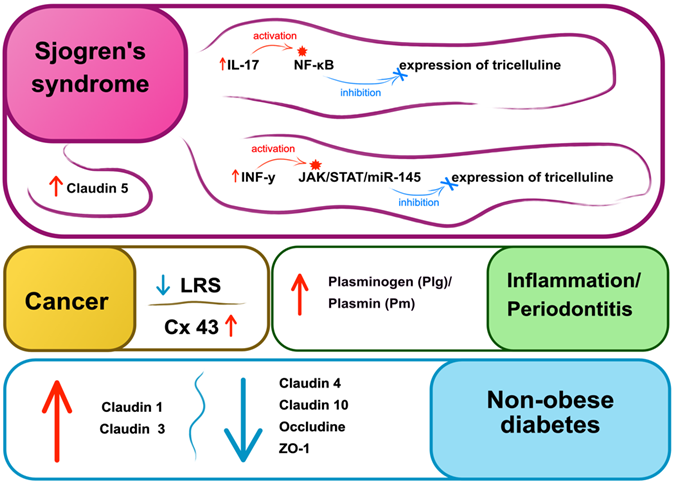
|
Figure 3.
Pathologies associated with damage to intercellular junctions of the salivary glands and the oral cavity epithelium. Cx-43 – connexin 43, LSR – lipolysis-stimulated lipoprotein receptor, ZO-1 – zonula occludens, NF-κB – nuclear factor kappa-light-chain-enhancer of activated B cells, INF-y – interferon gamma, JAK – Janus kinases, STAT – signal transducer and activator of transcription, miR-145 – microRNA-145.
|
Morphological changes of salivary glands and their intercellular contacts have been demonstrated in diabetes mellitus [125]. Ultrastructural damage, such as swollen mitochondria of the acini of the submandibular and parotid glands, as well as expansive endoplasmic reticulum and autophagosomes, were reported. Levels of claudin-1 (Cldn1) and claudin-3 (Cldn3) were increased, while levels of claudin-4 (Cldn4), occludin (Ocln), and ZO-1 were decreased in the submandibular salivary gland. High levels of Cldn1 and Cldn3 and lower levels of claudin-10 (Cldn10) and Ocln were demonstrated in the parotid gland. Such structural abnormalities of the submandibular and parotid salivary glands may contribute to hyposalivation [125]. Another study demonstrated that connexins 43 (Cx43) and connexins 26 (Cx26)were expressed in normal cells, whereas only Cx43 expression was observed in patients with early stage oral squamous cell carcinoma [126].
There are a number of contradictory studies regarding TJ expression and cancer progression [127]. Martin et al showed that loss of TJ could to tumor recurrence and carcinogenesis [128]. A number of other studies show that overexpression of TJ, namely JAM and claudins, were associated with tumor growth [129-131]. It can be assumed that depending on the stage of the disease and the type of cancer, increased or decreased expression of intercellular contact proteins contributes differently to the progression of carcinogenesis and tumor growth. Recently, a new observation showing that components of the plasminogen (Plg)/plasmin (Pm) system expressed in oral tissues, namely in the salivary gland tissues has been made. These components can contribute to microbial infection and inflammation, such as periodontitis [132].
Another structural component of the BSB is saliva. Saliva is a multifunctional fluid that lubricates the oral mucosa and contains electrolytes, antibacterial compounds, and a wide variety of enzymes involved in protecting the oral mucosa [133]. An interesting fact is that, despite its fluidity, saliva is not a Newtonian fluid, meaning that its viscosity changes with changes in the flow rate. This means that with changes in shear (flow rate), the viscosity of saliva changes; that is, with high shear, saliva is more fluid, while with low shear, saliva is more viscous. In other words, saliva is a viscoelastic material that exhibits both elastic (solid-like) and viscous (liquid-like) properties and reactions [134-135]. Saliva also exhibits thixotropic properties when structural changes in saliva occur under shear and are restored at rest [136-137]. In addition to time and shear, the rheological properties of saliva are also influenced by biochemical parameters such as Ca2+ concentration, total protein, and glycoproteins (mucins). Mucins have been shown to consist of 50-80% carbohydrates and are essential for the viscous properties of saliva. In one experiment, removal of mucins resulted in a decrease in saliva viscosity to the level of water. Thus, large, highly glycosylated glycoproteins with viscoelastic and protective properties play a crucial role in the viscous lubrication of the oral mucosa and are among the main factors that determine the rheological properties of saliva [138-139].
From a BSB perspective, blood supply and innervation are important for understanding the causes of changes in the composition of secreted saliva.
The vasculature of the salivary glands and the epithelium of the oral mucosa are represented by numerous arteries and veins and have a complex structure. It is worth noting that the volume of saliva produced per day is 1–2 liters, which is equal to the amount of urine produced in a physiologically normal human being and is directly dependent on arterial blood flow and vascular pressure [24]. The volume of the vascular network, the caliber of the vessels, and the pressure within them are of particular interest, as these characteristics will largely determine the rate of secretion, the volume of saliva secreted the condition of the salivary glands, and the epithelium of the oral mucosa [24].
The type of innervation directly influences the composition of secreted saliva. Thus, parasympathetic secretory innervation affects vasodilation of blood vessels, thereby increasing blood supply and, through acetylcholine and substance P, stimulates the secretion of serous saliva. Sympathetic nerves respond to norepinephrine, which causes the secretion of mucous saliva [140-141]. Nerve endings of nerve fibers are localized near the acinar cells of the salivary glands, duct cells and myoepithelial cells, and can be either naked or surrounded by Schwann cells. Parasympathetic and sympathetic innervation can have a direct effect on secretory cells, leading to an increase in intracellular Ca2+ levels, changes in cell membrane permeability, and the secretion of organic molecules, electrolytes, water and mucus in saliva [142]. In addition, nerve stimuli act on myoepithelial cells surrounding the acini and duct cells, which are regulated by both cholinergic and adrenergic neurotransmitters. This facilitates degranulation and fluid movement into the lumens of the salivary glands [143]. It has been shown that blockade of adrenergic receptors on myoepithelial cells leads to a slowdown in the fluid movement in the salivary glands without changing its secretion [144-146]. Steroidal and nonsteroidal hormones can be allosteric regulators of muscarinic receptors. Membrane cholesterol is able to bind and modulate the function of several G-protein-coupled receptors, including muscarinic acetylcholine receptors. This represents a new pharmacological approach to treatment of diseases associated with altered cholinergic signaling [147].
The oral immune system consists of three main compartments: the epithelial layer, the lamina propria, and the mucosa-associated lymphoid tissue (MALT) [148]. Pattern recognition receptors (PRRs) are immune cell proteins that detect and bind conserved pathogen-associated molecular patterns (PAMPs) and damage-associated molecules (DAMPs), thereby triggering the innate immune response. Currently identified PRR families include Toll-like receptors (TLRs), C-type lectin receptors (CLRs), nucleotide oligomerization domain-like receptors (NLRs), retinoic acid-inducible gene I-like receptors (RLRs), and AIM2-like receptors (ALRs) [148]. Furthermore, innate defense plays a key role in the activation and regulation of adaptive immunity (Fig. 4).
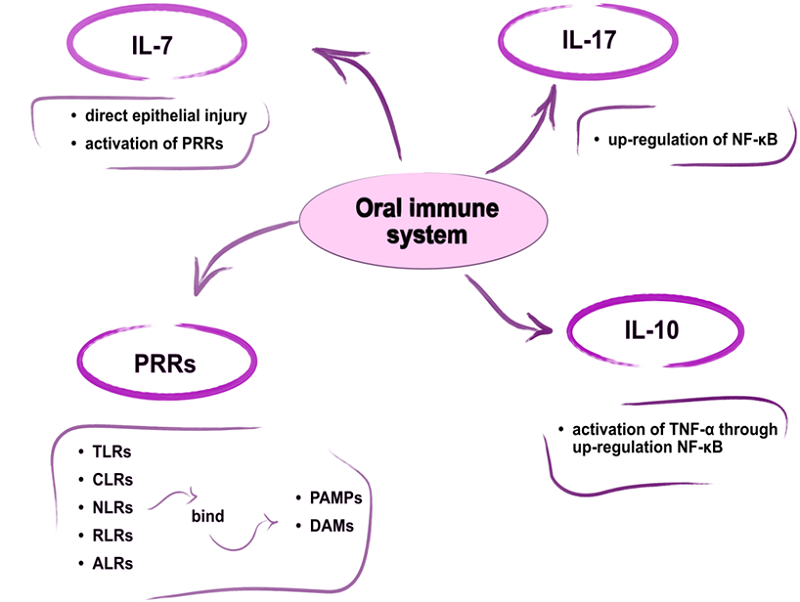
|
Figure 4.
Oral immune system. PRRs - pattern recognition receptors, PAMPs - pathogen-associated molecular patterns, DAMPs - damage-associated molecules, TNF-α - tumor necrosis factor-alpha, NF-κB - nuclear factor kappa-light-chain-enhancer of activated B cells, TLRs - Toll-like receptors, CLRs - C-type lectin receptors, NLRs - nucleotide oligomerization domain-like receptors, RLRs - retinoic acid-inducible gene I-like receptors, ALRs - AIM2 (Absent in melanoma 2)-like receptors.
|
The interleukin-7 cytokine-mediated immune pathway is induced within hours of epithelial cell injury or PRR activation [149]. Although IL-17 has been described as a T-cell-secreted cytokine, it has been shown that a significant portion of the IL-17 released during the inflammatory response is produced by innate immune cells [150]. Innate immune cell populations, which are an early source of IL-17 in response to stress, injury, or pathogens, are thought to reside in barrier tissues at the host-environment interface [150]. Epithelial cells are thought to perform a variety of sentinel functions, recognizing and mounting defenses against pathogens, in addition to their role as a physical barrier [150]. This region is also thought to be dominated by very strong immune tolerance, which allows for the monitoring and control of the interaction between the host's innate defenses and the microbiota [151]. IL-10 may play a protective role in head and neck squamous cell carcinoma. IL-10 has been shown to increase tumor necrosis factor-α (TNF-α) and lead to upregulation of NF-κB, which mediated the antitumor effect [152].
Due to their fundamental symbiotic lifestyle, the microbiota and innate immunity are ivolved in extensive two-way communication [153]. While the immune system influences and maintains the microbiota's microenvironment, the host microbiota restructures and promotes the immune system's tolerance of commensal and beneficial microbiota members [153]. Both the microbiota and the immune system also interact to mount necessary responses against pathogenic and harmful microorganisms. Achieving both local and systemic homeostasis while maintaining the host's biological integrity requires a highly regulated and coordinated interaction between the immune system and the microbiota. In this context, symbiosis and dysbiosis represent two extremes of the complex relationship between the oral microbial community and the immune responses to its presence. Many multifactorial disorders are thought to be dependent on and/or caused by alterations in the close interaction between the immune system and the microbiota [154].
In recent years, studies of microbiota metabolites and their interactions with the immune system have also opened a new dimension in understanding host-microbiota interactions. It is believed that host perception of bacterial metabolites is much more informative in determining the state of microbial colonization than recognition of microbial surface molecules, as metabolites provide information about the activity and functions of microorganisms, rather than their presence or absence [155-156].
The scientific community is presently engaged in rigorous research on the BSB, developing models that accurately replicate in vivo conditions. This review aims to enhance our understanding of the BSB by categorizing its components into core elements and those pertaining to the microenvironment. The core components consist of the salivary glands, oral epithelium, intercellular junction proteins responsible for barrier integrity and strength, saliva, and mucus. Microenvironmental components include innervation and blood vessels, local oral immunity, and the microbiome. Thus, the BSB is a broader concept, and its study should go beyond considering only the tissues located at the border between blood and saliva.
The authors declare that they received no external funding.
The authors declare that they have no conflicts of interest.
- Fröhlich, E. (2002) Aufbau und funktion von blut-gewebeschranken [Structure and function of blood-tissue barriers]. Dtsch Med Wochenschr, 127, 2629-2634. DOI
- Ruvinskaya, G.R., Mukhamedzhanova, L.R. (2013) Hematosalivary barrier: morphofunctional features in norm and pathology. Prakticheskaya Meditsina, 4, 21-25.
- Huykin, S.V., Akmalova, G.M. (2015). The concept of hematosalivary barrier. Meditsinskiy Vestnik Bashkortostana, 5, 103-107.
- Waespe, N., Strebel, S., Marino, D., Mattiello, V., Muet, F., Nava, T., Schindera, C., Belle, F.N., Mader, L., Spoerri, A., Kuehni, C.E., Ansari, M. (2021) Predictors for participation in DNA self-sampling of childhood cancer survivors in Switzerland. BMC Medical Research Methodology, 21(1), 236. DOI
- Lin, G.C., Friedl, H.P., Grabner, S., Gerhartl, A., Neuhaus, W. (2024) Transport of non-steroidal anti-inflammatory drugs across an oral mucosa epithelium in vitro model. Pharmaceutics, 16(4), 543. DOI
- Čižmárová, B., Tomečková, V., Hubková, B., Hurajtová, A., Ohlasová, J., Birková, A. (2022) Salivary redox homeostasis in human health and disease. International Journal of Molecular Sciences, 23(17), 10076. DOI
- Cong, X., Mao, X. D., Wu, L. L., Yu, G.Y. (2024) The role and mechanism of tight junctions in the regulation of salivary gland secretion. Oral Diseases, 30(1), 3–22. DOI
- Lin, G.C., Küng, E., Smajlhodzic, M., Domazet, S., Friedl, H.P., Angerer, J., Wisgrill, L., Berger, A., Bingle, L., Peham, J.R., Neuhaus, W. (2021) Directed transport of CRP across in vitro models of the BSB strengthens the feasibility of salivary CRP as biomarker for neonatal sepsis. Pharmaceutics, 13(2), 256. DOI
- Thaler, J., Tripisciano, C., Nieuwland, R. (2024) Investigations on the hemostatic potential of physiological body fluids. Hamostaseologie, 44, 377-385. DOI
- Lin, G.C., Tevini, J., Mair, L., Friedl, H.P., Fuchs, D., Felder, T., Gostner, J.M., Neuhaus, W. (2024) Investigations towards tryptophan uptake and transport across an in vitro model of the oral mucosa epithelium. International Journal of Tryptophan Research, 17, 11786469241266312. DOI
- Gonçalves, A., Ambrósio, A.F., Fernandes, R. (2013) Regulation of claudins in blood-tissue barriers under physiological and pathological states. Tissue Barriers, 1(3), e24782. DOI
- Wen, Q., Tang, E.I., Gao, Y., Jesus, T.T., Chu, D.S., Lee, W.M., Wong, C.K.C., Liu, Y.X., Xiao, X., Silvestrini, B., Cheng, C.Y. (2018) Signaling pathways regulating blood-tissue barriers - Lesson from the testis. Biochimica et Biophysica Acta. Biomembranes, 1860(1), 141–153. DOI
- Hyvärinen, E., Savolainen, M., Mikkonen, J.J.W., Kullaa, A.M. (2021) Salivary metabolomics for diagnosis and monitoring diseases: challenges and possibilities. Metabolites, 11(9), 587. DOI
- Cui, Y., Yang, M., Zhu, J., Zhang, H., Duan, Z., Wang, S., Liao, Z., Liu, W. (2022) Developments in diagnostic applications of saliva in human organ diseases. Medicine in Novel Technology and Devices, 13, 100115. DOI
- Boroumand, M., Olianas, A., Cabras, T., Manconi, B., Fanni, D., Faa, G., Desiderio, C., Messana, I., Castagnola, M. (2021) Saliva, a bodily fluid with recognized and potential diagnostic applications. Journal of Separation Science, 44(19), 3677–3690. DOI
- Gartner, L.P. (1994) Oral anatomy and tissue types. Seminars in Dermatology, 13.
- Şenel, S. (2021) An overview of physical, microbiological and immune barriers of oral mucosa. International Journal of Molecular Sciences, 22(15), 7821. DOI
- Garant, P.R. (2003) Oral cells and tissues. New York State Dental Journal, 60.
- Wertz, P.W. (2021) Roles of lipids in the permeability barriers of skin and oral mucosa. International Journal of Molecular Sciences, 22(10), 5229. DOI
- Patel, V.N., Hoffman, M.P. (2014) Salivary gland development: a template for regeneration. Seminars in Cell Developmental Biology, 25-26, 52–60. DOI
- Lin, G.C., Smajlhodzic, M., Bandian, A.M., Friedl, H.P., Leitgeb, T., Oerter, S., Stadler, K., Giese, U., Peham, J.R., Bingle, L., Neuhaus, W. (2020) An in vitro barrier model of the human submandibular salivary gland epithelium based on a single cell clone of cell line HTB- 41: establishment and application for biomarker transport studies. Biomedicines, 8(9), 302. DOI
- Punj, A. (2019) Secretions of human salivary gland. In: Salivary Glands: New Approaches in Diagnostics and Treatment, edited by Güvenç, I.A. London: InTechOpen (Chapter 4).
- Kim, S.K., Nasjleti, C.E., Han, S.S. (1972) The secretion processes in mucous and serous secretory cells of the rat sublingual gland. Journal of Ultrastructure Research, 38(3), 371–389. DOI
- Holmberg, K.V., Hoffman, M.P. (2014) Anatomy, biogenesis and regeneration of salivary glands. Monographs in Oral Science, 24, 1–13. DOI
- Baker, O.J. (2016) Current trends in salivary gland tight junctions. Tissue Barriers, 4(3), e1162348. DOI
- de Paula, F., Teshima, T.H.N., Hsieh, R., Souza, M.M., Coutinho- Camillo, C.M., Nico, M.M.S., Lourenco, S.V. (2017) The expression of water channel proteins during human salivary gland development: a topographic study of aquaporins 1, 3 and 5. Journal of Molecular Histology, 48(5-6), 329–336. DOI
- Larsen, H.S., Aure, M.H., Peters, S.B., Larsen, M., Messelt, E.B., Kanli Galtung, H. (2011) Localization of AQP5 during development of the mouse submandibular salivary gland. Journal of Molecular Histology, 42(1), 71–81. DOI
- Hsieh, M.S., Jeng, Y.M., Lee, Y.H. (2019) Mist1: a novel nuclear marker for acinic cell carcinoma of the salivary gland. Virchows Archiv, 475(5), 617–624. DOI
- Pin, C.L., Bonvissuto, A.C., Konieczny, S.F. (2000) Mist1 expression is a common link among serous exocrine cells exhibiting regulated exocytosis. The Anatomical Record, 259(2), 157–167. DOI
- Valstar, M.H., de Bakker, B.S., Steenbakkers, R.J.H. M., de Jong, K.H., Smit, L.A., Klein Nulent, T.J.W., van Es, R.J.J., Hofland, I., de Keizer, B., Jasperse, B., Balm, A.J.M., van der Schaaf, A., Langendijk, J.A., Smeele, L.E., Vogel, W.V. (2021) The tubarial salivary glands: A potential new organ at risk for radiotherapy. Radiotherapy and oncology: journal of the European Society for Therapeutic Radiology and Oncology, 154, 292–298. DOI
- Dave, M. (2020) New salivary gland organs discovered. British Dental Journal, 229(9), 573. DOI
- Suvvari, T.K., Arigapudi, N. (2020) The tubarial glands: Discovered but not defined–A narrative review. Journal of Radiation and Cancer Research, 11, 140-141. DOI
- Singh, V., Reddy, K.C. (2021) Discovery of new organs in human throat: The tubarial salivary glands. Journal of the Anatomical Society of India, 70, 1-2. DOI
- Ebrahim, A., Reich, C., Wilde, K., Salim, A. M., Hyrcza, M.D., Willetts, L. (2025) A comprehensive analysis of the tubarial glands. Anatomical Record, 308(5), 1425–1437. DOI
- Zhu, W., Wang, Y., Guan, Y., Lu, Y., Li, Y., Sun, L., Wang, Y. (2024) Rapamycin can alleviate the submandibular gland pathology of Sjögren’s syndrome by limiting the activation of cGAS–STING signaling pathway. Inflammopharmacology, 32, 1113-31. DOI
- Sisto, M., Ribatti, D., Lisi, S. (2021) SMADS-mediate molecular mechanisms in Sjögren’s syndrome. International Journal of Molecular Sciences, 22(6), 3203. DOI
- Fouani, M., Basset, C. A., Jurjus, A. R., Leone, L.G., Tomasello, G., Leone, A. (2021) Salivary gland proteins alterations in the diabetic milieu. Journal of Molecular Histology, 52(5), 893–904. DOI
- Genco, R.J., Borgnakke, W.S. (2020) Diabetes as a potential risk for periodontitis: association studies. Periodontol, 2000, 83, 40-45. DOI
- Chen, S.Y., Wang, Y., Zhang, C.L., Yang, Z.M. (2020) Decreased basal and stimulated salivary parameters by histopathological lesions and secretory dysfunction of parotid and submandibular glands in rats with type 2 diabetes. Experimental and Therapeutic Medicine, 19(4), 2707–2719. DOI
- Klein Hesselink, E.N., Brouwers, A.H., de Jong, J.R., van der Horst-Schrivers, A.N., Coppes, R.P., Lefrandt, J.D., Jager, P.L., Vissink, A., Links, T.P. (2016) Effects of radioiodine treatment on salivary gland function in patients with differentiated thyroid carcinoma: a prospective study. Journal of Nuclear Medicine, 57(11), 1685–1691. DOI
- Jung, J.H., Lee, C.H., Son, S.H., Jeong, J.H., Jeong, S.Y., Lee, S.W., Lee, J., Ahn, B.C. (2017) High prevalence of thyroid disease and role of salivary gland scintigraphy in patients with xerostomia. Nuclear Medicine and Molecular Imaging, 51(2), 169–177. DOI
- Abetz, L.M., Savage, N.W. (2009) Burning mouth syndrome and psychological disorders. Australian Dental Journal, 54(2), 84–173. DOI
- Verhoeff, M.C., Koutris, M., Vries, R., Berendse, H.W., Dijk, K.D.V., Lobbezoo, F. (2023) Salivation in Parkinson’s disease: A scoping review. Gerodontology, 40(1), 26–38. DOI
- Jääskeläinen, S.K. (2018) Is burning mouth syndrome a neuropathic pain condition? Pain, 159(3), 610–613. DOI
- Sørensen, C.E., Hansen, N.L., Mortensen, E.L., Lauritzen, M., Osler, M., Pedersen, A.M. L. (2018) Hyposalivation and poor dental health status are potential correlates of age-related cognitive decline in late midlife in Danish men. Frontiers in Aging Neuroscience, 10, 10. DOI
- Yuan, Y., Jiao, B., Qu, L., Yang, D., Liu, R. (2023) The development of COVID-19 treatment. Frontiers in Immunology, 14, 1125246. DOI
- Meer, S. (2019) Human immunodeficiency virus and salivary gland pathology: an update. Oral Surgery, Oral Medicine, Oral Pathology and Oral Radiology, 128(1), 52–59. DOI
- Domingues, N.B., Mariusso, M.R., Tanaka, M.H., Scarel- Caminaga, R.M., Mayer, M.P.A., Brighenti, F.L., Zuanon, Â.C.C., Ibuki, F.K., Nogueira, F.N., Giro, E.M.A. (2017) Reduced salivary flow rate and high levels of oxidative stress in whole saliva of children with Down syndrome. Special Care in Dentistry, 37(6), 269–276. DOI
- Seymen, F., Koruyucu, M., Toptanci, I. R., Balsak, S., Dedeoglu, S., Celepkolu, T., Shin, T.J., Hyun, H.K., Kim, Y.J., Kim, J.W. (2017) Novel FGF10 mutation in autosomal dominant aplasia of lacrimal and salivary glands. Clinical Oral Investigations, 21(1), 167–172. DOI
- Saeves, R., Reseland, J.E., Kvam, B.M., Sandvik, L., Nordgarden, H. (2012) Saliva in Prader-Willi syndrome: quantitative and qualitative characteristics. Archives of Oral Biology, 57(10), 1335–1341. DOI
- Vasconcelos, G., Stenehjem, J.S., Axelsson, S., Saeves, R. (2022) Craniofacial and dentoalveolar morphology in individuals with Prader-Willi syndrome: a case-control study. Orphanet Journal of Rare Diseases, 17(1), 77. DOI
- Ramírez, L., Sánchez, I., Muñoz, M., Martínez-Acitores, M.L., Garrido, E., Hernández, G., López-Pintor, R.M. (2023) Risk factors associated with xerostomia and reduced salivary flow in hypertensive patients. Oral Diseases, 29(3), 1299–1311. DOI
- Drent, M., Crouser, E. D., Grunewald, J. (2021) Challenges of Sarcoidosis and Its Management. The New England Journal of Medicine, 385(11), 1018–1032. DOI
- Arany, S., Kopycka-Kedzierawski, D.T., Caprio, T.V., Watson, G.E. (2021) Anticholinergic medication: Related dry mouth and effects on the salivary glands. Oral Surgery, Oral Medicine, Oral Pathology and Oral Radiology, 132(6), 662–670. DOI
- Toan, N.K., Tai, N. C., Kim, S.A., Ahn, S.G. (2021) Choline acetyltransferase induces the functional regeneration of the salivary gland in aging SAMP1/Kl-/-Mice. International Journal of Molecular Sciences, 22(1), 404. DOI
- Song, W., Liu, H., Su, Y., Zhao, Q., Wang, X., Cheng, P., Wang, H. (2024) Current developments and opportunities of pluripotent stem cells-based therapies for salivary gland hypofunction. Frontiers in Cell and Developmental Biology, 12, 1346996. DOI
- Roussa, E. (2011) Channels and transporters in salivary glands. Cell and Tissue Research, 343(2), 263–287. DOI
- Catalán, M.A., Nakamoto, T., Melvin, J.E. (2009) The salivary gland fluid secretion mechanism. The Journal of Medical Investigation: JMI, 56 Suppl, 192–196. DOI
- Lapczuk-Romanska, J., Busch, D., Gieruszczak, E., Drozdzik, A., Piotrowska, K., Kowalczyk, R., Oswald, S., Drozdzik, M. (2019) Membrane transporters in human parotid gland-targeted proteomics approach. International Journal of Molecular Sciences, 20(19), 4825. DOI
- Sun, Q. F., Sun, Q. H., Du, J., Wang, S. (2008) Differential gene expression profiles of normal human parotid and submandibular glands. Oral Diseases, 14(6), 500–509. DOI
- Petersen, A.M., Small, C.M., Yan, Y.L., Wilson, C., Batzel, P., Bremiller, R.A., Buck, C.L., von Hippel, F.A., Cresko, W.A., Postlethwait, J.H. (2022) Evolution and developmental expression of the sodium-iodide symporter (NIS, slc5a5) gene family: Implications for perchlorate toxicology. Evolutionary Applications, 15(7), 1079– 1098. DOI
- Skrypnyk, M., Yatsenko, T., Riabets, O., Zuieva, O., Rodionova, I., Skikevych, M., Salama, Y., Osada, T., Tobita, M., Takahashi, S., Hattori, N., Takahashi, K., Hattori, K., Heissig, B. (2025) Potassium iodide induces apoptosis in salivary gland cancer cells. International Journal of Molecular Sciences, 26, 5199. DOI
- Angelow, S., Ahlstrom, R., Yu, A.S. (2008) Biology of claudins. American journal of physiology. Renal Physiology, 295(4), F867– F876. DOI
- Furuse, M., Tsukita, S. (2006). Claudins in occluding junctions of humans and flies. Trends in Cell Biology, 16(4), 181–188. https://doi. org/10.1016/j.tcb.2006.02.006
- Ikenouchi, J., Furuse, M., Furuse, K., Sasaki, H., Tsukita, S., Tsukita, S. (2005) Tricellulin constitutes a novel barrier at tricellular contacts of epithelial cells. The Journal of Cell Biology, 171(6), 939–945. DOI
- Do, T.T., Nguyen, V.T., Nguyen, N.T.N., Duong, K.T.T., Nguyen, T.T.M., Le, D.N.T., Nguyen, T.H. (2024) A review of a breakdown in the barrier: tight junction dysfunction in dental diseases. Clinical, Cosmetic and Investigational Dentistry, 16, 513–531. DOI
- Hirabayashi, S., Tajima, M., Yao, I., Nishimura, W., Mori, H., Hata, Y. (2003) JAM4, a junctional cell adhesion molecule interacting with a tight junction protein, MAGI-1. Molecular and Cellular Biology, 23(12), 4267–4282. DOI
- Nasdala, I., Wolburg-Buchholz, K., Wolburg, H., Kuhn, A., Ebnet, K., Brachtendorf, G., Samulowitz, U., Kuster, B., Engelhardt, B., Vestweber, D., Butz, S. (2002) A transmembrane tight junction protein selectively expressed on endothelial cells and platelets. The Journal of Biological Chemistry, 277(18), 16294–16303. DOI
- Monteiro, A.C., Sumagin, R., Rankin, C.R., Leoni, G., Mina, M.J., Reiter, D.M., Stehle, T., Dermody, T.S., Schaefer, S.A., Hall, R.A., Nusrat, A., Parkos, C.A. (2013) JAM-A associates with ZO-2, afadin, and PDZ-GEF1 to activate Rap2c and regulate epithelial barrier function. Molecular Biology of the Cell, 24(18), 2849–2860. DOI
- Ebnet, K., Schulz, C.U., Meyer Zu Brickwedde, M.K., Pendl, G.G., Vestweber, D. (2000) Junctional adhesion molecule interacts with the PDZ domain-containing proteins AF-6 and ZO-1. The Journal of Biological Chemistry, 275(36), 27979–27988. DOI
- Gow, A., Davies, C., Southwood, C.M., Frolenkov, G., Chrustowski, M., Ng, L., Yamauchi, D., Marcus, D.C., Kachar, B. (2004) Deafness in Claudin 11-null mice reveals the critical contribution of basal cell tight junctions to stria vascularis function. The Journal of Neuroscience, 24(32), 7051–7062. DOI
- Tsukita, S., Furuse, M. (2002) Claudin-based barrier in simple and stratified cellular sheets. Current Opinion in Cell Biology, 14(5), 531–536. DOI
- Furuse, M., Hata, M., Furuse, K., Yoshida, Y., Haratake, A., Sugitani, Y., Noda, T., Kubo, A., Tsukita, S. (2002) Claudin-based tight junctions are crucial for the mammalian epidermal barrier: a lesson from claudin-1-deficient mice. The Journal of Cell Biology, 156(6), 1099–1111. DOI
- Furuse, M. (2009) Knockout animals and natural mutations as experimental and diagnostic tool for studying tight junction functions in vivo. Biochimica et Biophysica Acta, 1788(4), 813–819. DOI
- Saitou, M., Furuse, M., Sasaki, H., Schulzke, J.D., Fromm, M., Takano, H., Noda, T., Tsukita, S. (2000) Complex phenotype of mice lacking occludin, a component of tight junction strands. Molecular Biology of the Cell, 11(12), 4131–4142. DOI
- Shen, L., Turner, J.R. (2005) Actin depolymerization disrupts tight junctions via caveolae-mediated endocytosis. Molecular Biology of the Cell, 16(9), 3919–3936. DOI
- Yu, A.S., McCarthy, K.M., Francis, S.A., McCormack, J.M., Lai, J., Rogers, R.A., Lynch, R.D., Schneeberger, E.E. (2005) Knockdown of occludin expression leads to diverse phenotypic alterations in epithelial cells. American Journal of Physiology. Cell Physiology, 288(6), C1231–C1241. DOI
- Utech, M., Mennigen, R., Bruewer, M. (2010) Endocytosis and recycling of tight junction proteins in inflammation. Journal of Biomedicine Biotechnology, 2010, 484987. DOI
- Chen, Y.H., Lu, Q., Goodenough, D.A., Jeansonne, B. (2002) Nonreceptor tyrosine kinase c-Yes interacts with occludin during tight junction formation in canine kidney epithelial cells. Molecular Biology of the Cell, 13(4), 1227–1237. DOI
- Seth, A., Sheth, P., Elias, B. C., Rao, R. (2007) Protein phosphatases 2A and 1 interact with occludin and negatively regulate the assembly of tight junctions in the CACO-2 cell monolayer. The Journal of Biological Chemistry, 282(15), 11487–11498. DOI
- Furuse, M., Hirase, T., Itoh, M., Nagafuchi, A., Yonemura, S., Tsukita, S., Tsukita, S. (1993) Occludin: a novel integral membrane protein localizing at tight junctions. The Journal of Cell Biology, 123(6 Pt 2), 1777–1788. DOI
- Furuse, M. (2010) Molecular basis of the core structure of tight junctions. Cold Spring Harbor Perspectives in Biology, 2(1), a002907. DOI
- Ikenouchi, J., Sasaki, H., Tsukita, S., Furuse, M., Tsukita, S. (2008) Loss of occludin affects tricellular localization of tricellulin. Molecular Biology of the Cell, 19(11), 4687–4693. DOI
- Krug, S.M., Amasheh, S., Richter, J.F., Milatz, S., Günzel, D., Westphal, J.K., Huber, O., Schulzke, J.D., Fromm, M. (2009) Tricellulin forms a barrier to macromolecules in tricellular tight junctions without affecting ion permeability. Molecular Biology of the Cell, 20(16), 3713–3724. DOI
- Brummelkamp, T.R., Bernards, R., Agami, R. (2002) A system for stable expression of short interfering RNAs in mammalian cells. Science (New York, N.Y.), 296(5567), 550–553. DOI
- Riazuddin, S., Ahmed, Z.M., Fanning, A.S., Lagziel, A., Kitajiri, S., Ramzan, K., Khan, S.N., Chattaraj, P., Friedman, P.L., Anderson, J.M., Belyantseva, I.A., Forge, A., Riazuddin, S., Friedman, T.B. (2006) Tricellulin is a tight-junction protein necessary for hearing. American Journal of Human Genetics, 79(6), 1040–1051. DOI
- Umeda, K., Ikenouchi, J., Katahira-Tayama, S., Furuse, K., Sasaki, H., Nakayama, M., Matsui, T., Tsukita, S., Furuse, M., Tsukita, S. (2006) ZO-1 and ZO-2 independently determine where claudins are polymerized in tight-junction strand formation. Cell, 126(4), 741–754. DOI
- Yamazaki, Y., Umeda, K., Wada, M., Nada, S., Okada, M., Tsukita, S., Tsukita, S. (2008) ZO-1- and ZO-2-dependent integration of myosin-2 to epithelial zonula adherens. Molecular Biology of the Cell, 19(9), 3801–3811. DOI
- Ikenouchi, J., Umeda, K., Tsukita, S., Furuse, M., Tsukita, S. (2007) Requirement of ZO-1 for the formation of belt-like adherens junctions during epithelial cell polarization. The Journal of Cell Biology, 176(6), 779–786. DOI
- Umeda, K., Matsui, T., Nakayama, M., Furuse, K., Sasaki, H., Furuse, M., Tsukita, S. (2004) Establishment and characterization of cultured epithelial cells lacking expression of ZO-1. The Journal of Biological Chemistry, 279(43), 44785–44794. DOI
- McNeil, E., Capaldo, C. T., Macara, I. G. (2006) Zonula occludens-1 function in the assembly of tight junctions in Madin- Darby canine kidney epithelial cells. Molecular Biology of the Cell, 17(4), 1922–1932. DOI
- Pinto-Dueñas, D.C., Hernández-Guzmán, C., Marsch, P.M., Wadurkar, A.S., Martín-Tapia, D., Alarcón, L., Vázquez-Victorio, G., Méndez-Méndez, J.V., Chanona-Pérez, J.J., Nangia, S., González- Mariscal, L. (2024) The role of ZO-2 in modulating JAM-A and γ-actin junctional recruitment, apical membrane and tight junction tension, and cell response to substrate stiffness and topography. International Journal of Molecular Sciences, 25(5), 2453. DOI
- Haskins, J., Gu, L., Wittchen, E.S., Hibbard, J., Stevenson, B.R. (1998) ZO-3, a novel member of the MAGUK protein family found at the tight junction, interacts with ZO-1 and occludin. The Journal of Cell Biology, 141(1), 199–208. DOI
- Liu, W., Cui, Y., Wei, J., Sun, J., Zheng, L., Xie, J. (2020) Gap junction-mediated cell-to-cell communication in oral development and oral diseases: a concise review of research progress. International Journal of Oral Science, 12(1), 17. DOI
- Krutovskikh, V., Yamasaki, H. (2000) Connexin gene mutations in human genetic diseases. Mutation Research, 462(2-3), 197–207. DOI
- Beyer, E.C., Berthoud, V.M. (2018) Gap junction gene and protein families: Connexins, innexins, and pannexins. Biochimica et Biophysica Acta. Biomembranes, 1860(1), 5–8. DOI
- Rodríguez-Sinovas, A., Ruiz-Meana, M., Denuc, A., García-Dorado, D. (2018) Mitochondrial Cx43, an important component of cardiac preconditioning. Biochimica et Biophysica Acta. Biomembranes, 1860(1), 174–181. DOI
- Zappitelli, T., Aubin, J.E. (2014) The “connexin” between bone cells and skeletal functions. Journal of Cellular Biochemistry, 115, 1646-1658. DOI
- Epifantseva, I., Shaw, R.M. (2018) Intracellular trafficking pathways of Cx43 gap junction channels. Biochimica et Biophysica Acta. Biomembranes, 1860(1), 40–47. DOI
- Pereda, A.E., Curti, S., Hoge, G., Cachope, R., Flores, C.E., Rash, J.E. (2013) Gap junction-mediated electrical transmission: regulatory mechanisms and plasticity. Biochimica et Biophysica Acta, 1828(1), 134–146. DOI
- Hervé, J.C., Derangeon, M. (2013) Gap-junction-mediated cellto- cell communication. Cell and Tissue Research, 352(1), 21–31. DOI
- Dupont, G., Combettes, L., Leybaert, L. (2007) Calcium dynamics: spatio-temporal organization from the subcellular to the organ level. International Review of Cytology, 261, 193–245. DOI
- Li, Y.Q., Tan, S.S., Wu, D., Zhang, Q., Wang, T., Zheng, G. (2025) The role of intracellular and extracellular copper compartmentalization in Alzheimer’s disease pathology and its implications for diagnosis and therapy. Frontiers in Neuroscience, 19, 1553064. DOI
- Al-Amoudi, A., Castaño-Diez, D., Devos, D.P., Russell, R.B., Johnson, G.T., Frangakis, A.S. (2011) The three-dimensional molecular structure of the desmosomal plaque. Proceedings of the National Academy of Sciences of the United States of America, 108(16), 6480–6485. DOI
- He, Y., Xu, M., Ouyang, J., Zhao, L., Ma, T., Zhang, X., Wang, R., Shang, H., Liang, G. (2025) Keratin-72 restricts HIV-1 infection in resting CD4+ T cells by sequestering capsids in intermediate filaments. Nature Communications, 16(1), 2998. DOI
- Parry, D.A.D. (2021) Structures of the ß-Keratin filaments and keratin intermediate filaments in the epidermal appendages of birds and reptiles (Sauropsids). Genes, 12(4), 591. DOI
- Louka, P., Kyriakou, C., Diakourti, I., Skourides, P. (2025) Plakophilin 3 is involved in basal body docking in multiciliated cells. International Journal of Molecular Sciences, 26(11), 5381. DOI
- Bass-Zubek, A.E., Godsel, L.M., Delmar, M., Green, K.J. (2009) Plakophilins: multifunctional scaffolds for adhesion and signaling. Current Opinion in Cell Biology, 21(5), 708–716. DOI
- Zhang, J., Gutierrez-Lara, E.J., Do, A., Nguyen, L., Nair, A., Selvan, N., Fenn, T., Adler, E., Khanna, R., Sheikh, F. (2025) Preclinical efficacy and safety of AAVrh10-based plakophilin-2 gene therapy (LX2020) as a treatment for arrhythmogenic cardiomyopathy. NPJ Regenerative Medicine, 10(1), 17. DOI
- Zou, Y., Lu, J., Lian, Z., Jia, J., Shen, J., Li, Q., Wong, J. M. J., Jin, K., Yan, W., Ren, X., Zhang, Y., Huang, C., Yang, H., Huang, F., Li, J., Zhai, J., Xu, Y., Xu, X., Yu, H., Jin, Y., Dai, Y. (2025) Modified mRNA treatment restores cardiac function in desmocollin- 2-deficient mouse models of arrhythmogenic right ventricular cardiomyopathy. Circulation, 151(25), 1780–1796. DOI
- Khan, K., Hardy, R., Haq, A., Ogunbiyi, O., Morton, D., Chidgey, M. (2006) Desmocollin switching in colorectal cancer. British Journal of Cancer, 95(10), 1367–1370. DOI
- Elias, P.M., Matsuyoshi, N., Wu, H., Lin, C., Wang, Z.H., Brown, B.E., Stanley, J.R. (2001) Desmoglein isoform distribution affects stratum corneum structure and function. The Journal of Cell Biology, 153(2), 243–249. DOI
- Yuan, Z.Y., Cheng, L.T., Wang, Z.F., Wu, Y.Q. (2021) Desmoplakin and clinical manifestations of desmoplakin cardiomyopathy. Chinese Medical Journal, 134(15), 1771–1779. DOI
- Choi, H. J., Weis, W. I. (2016) Purification and Structural Analysis of Desmoplakin. Methods in Enzymology, 569, 197–213. DOI
- Lialios, P., Alimperti, S. (2025) Role of E-cadherin in epithelial barrier dysfunction: implications for bacterial infection, inflammation, and disease pathogenesis. Frontiers in Cellular and Infection Microbiology, 15, 1506636. DOI
- Li, Y., Altorelli, N.L., Bahna, F., Honig, B., Shapiro, L., Palmer, A.G., 3rd (2013) Mechanism of E-cadherin dimerization probed by NMR relaxation dispersion. Proceedings of the National Academy of Sciences of the United States of America, 110(41), 16462–16467. DOI
- Kobecki, J., Gajdzis, P., Mazur, G., Chabowski, M. (2022) Nectins and nectin-like molecules in colorectal cancer: role in diagnostics, prognostic values, and emerging treatment options: a literature review. Diagnostics (Basel, Switzerland), 12(12), 3076. DOI
- Harrison, O.J., Vendome, J., Brasch, J., Jin, X., Hong, S., Katsamba, P.S., Ahlsen, G., Troyanovsky, R.B., Troyanovsky, S.M., Honig, B., Shapiro, L. (2012) Nectin ectodomain structures reveal a canonical adhesive interface. Nature Structural Molecular Biology, 19(9), 906–915. DOI
- Mao, X., Li, H., Min, S., Su, J., Wei, P., Zhang, Y., He, Q., Wu, L., Yu, G., Cong, X. (2025) Loss of tricellular tight junction tricellulin leads to hyposalivation in Sjögren’s syndrome. International Journal of Oral Science, 17, 22. DOI
- Zhang, L.W., Cong, X., Zhang, Y., Wei, T., Su, Y.C., Serrão, A.C., Brito, A.R., Jr, Yu, G. Y., Hua, H., Wu, L. L. (2016) Interleukin-17 impairs salivary tight junction integrity in Sjögren’s syndrome. Journal of Dental Research, 95(7), 784–792. DOI
- Cong, X., Zhang, X.M., Zhang, Y., Wei, T., He, Q.H., Zhang, L.W., Hua, H., Lee, S.W., Park, K., Yu, G. Y., Wu, L.L. (2018) Disruption of endothelial barrier function is linked with hyposecretion and lymphocytic infiltration in salivary glands of Sjögren’s syndrome. Biochimica et Biophysica Acta, 1864(10), 3154–3163. DOI
- Lisi, S., Sisto, M., D’Amore, M., Lofrumento, D.D., Ribatti, D. (2013) Emerging avenues linking inflammation, angiogenesis and Sjögren’s syndrome. Cytokine, 61(3), 693–703. DOI
- McCall, A.D., Baker, O.J. (2015) Characterization of angiogenesis and lymphangiogenesis in human minor salivary glands with Sjögren’s syndrome. The Journal of Histochemistry and Cytochemistry, 63(5), 340–349. DOI
- Nishida, S., Konno, T., Kohno, T., Ohyanagi, M., Nakano, M., Ohwada, K., Obata, K., Kakuki, T., Kakiuchi, A., Kurose, M., Takano, K., Kojima, T. (2025) Treatment with TNFα and lipolysis-stimulated lipoprotein receptor (LSR) antibody in the presence of HDAC inhibitors promotes apoptosis in human salivary duct adenocarcinoma. Tissue Barriers, 13(3), 2437215. DOI
- Huang, Y., Mao, Q.Y., Shi, X.J., Cong, X., Zhang, Y., Wu, L.L., Yu, G. Y., Xiang, R.L. (2020) Disruption of tight junctions contributes to hyposalivation of salivary glands in a mouse model of type 2 diabetes. Journal of Anatomy, 237(3), 556–567. DOI
- Brockmeyer, P., Jung, K., Perske, C., Schliephake, H., Hemmerlein, B. (2014) Membrane connexin 43 acts as an independent prognostic marker in oral squamous cell carcinoma. International Journal of Oncology, 45(1), 273–281. DOI
- Samiei, M., Ahmadian, E., Eftekhari, A., Eghbal, M. A., Rezaie, F., Vinken, M. (2019) Cell junctions and oral health. EXCLI Journal, 18, 317–330. DOI
- Martin, T.A., Mansel, R.E., Jiang, W.G. (2010) Loss of occludin leads to the progression of human breast cancer. International Journal of Molecular Medicine, 26(5), 723–734. DOI
- Dos Reis, P.P., Bharadwaj, R.R., Machado, J., Macmillan, C., Pintilie, M., Sukhai, M.A., Perez-Ordonez, B., Gullane, P., Irish, J., K amel-Reid, S. (2008) Claudin 1 overexpression increases invasion and is associated with aggressive histological features in oral squamous cell carcinoma. Cancer, 113(11), 3169–3180. DOI
- Oku, N., Sasabe, E., Ueta, E., Yamamoto, T., Osaki, T. (2006) Tight junction protein claudin-1 enhances the invasive activity of oral squamous cell carcinoma cells by promoting cleavage of laminin-5 gamma2 chain via matrix metalloproteinase (MMP)-2 and membrane-type MMP-1. Cancer Research, 66(10), 5251–5257. DOI
- Leech, A.O., Cruz, R.G., Hill, A.D., Hopkins, A.M. (2015) Paradigms lost-an emerging role for over-expression of tight junction adhesion proteins in cancer pathogenesis. Annals of Translational Medicine, 3(13), 184. DOI
- Yatsenko, T., Skrypnyk, M., Troyanovska, O., Tobita, M., Osada, T., Takahashi, S., Hattori, K., Heissig, B. (2023) The role of the plasminogen/plasmin system in inflammation of the oral cavity. Cells, 12(3), 445. DOI
- Feller, L., Altini, M., Khammissa, R. A., Chandran, R., Bouckaert, M., Lemmer, J. (2013) Oral mucosal immunity. Oral Surgery, Oral Medicine, Oral Pathology and Oral Radiology, 116(5), 576–583. DOI
- Mishra, A.A., Almhöjd, U., Çevik-Aras, H., Fisic, A., Olofsson, R., Almståhl, A., Kádár, R. (2025) The complex shear time response of saliva in healthy individuals. Physics of Fluids, 37, 011911. DOI
- Dewhirst, F.E., Chen, T., Izard, J., Paster, B.J., Tanner, A.C., Yu, W.H., Lakshmanan, A., Wade, W.G. (2010) The human oral microbiome. Journal of Bacteriology, 192(19), 5002–5017. DOI
- Larson, R.G., Wei, Y. (2019) A review of thixotropy and its rheological modeling. Journal of Rheology, 63, 477–501. DOI
- Bugarin-Castillo, Y., Bou-Fadel, P., Mohamed-Ismail, S., Huang, N., Saint-Eve, A., Mathieu, V., Ramaioli, M. (2024) On the rheological and sensory properties of a novel natural salivary substitute. European Journal of Pharmaceutical Sciences, 199, 106802. DOI
- Phelps, C.F. (1978) Biosynthesis of mucus glycoprotein. British medical bulletin, 34(1), 43–48. DOI
- Veerman, E.C., Valentijn-Benz, M., Nieuw Amerongen, A.V. (1989) Viscosity of human salivary mucins: effect of pH and ionic strength and role of sialic acid. Journal de Biologie Buccale, 17(4), 297–306.
- Ekström, J. (1989) Autonomic control of salivary secretion. Proceedings of the Finnish Dental Society, 85(4-5), 323–363.
- Sato, T., Mito, K., Ishii, H. (2020) Relationship between impaired parasympathetic vasodilation and hyposalivation in parotid glands associated with type 2 diabetes mellitus. American Journal of Physiology, 318(5), R940–R949. DOI
- Ferreira, J.N., Hoffman, M.P. (2013) Interactions between developing nerves and salivary glands. Organogenesis, 9(3), 199–205. DOI
- Ozdemir, T., Srinivasan, P.P., Zakheim, D.R., Harrington, D.A., Witt, R.L., Farach-Carson, M.C., Jia, X., Pradhan-Bhatt, S. (2017) Bottom-up assembly of salivary gland microtissues for assessing myoepithelial cell function. Biomaterials, 142, 124–135. DOI
- Schneyer, C.A., Humphreys-Beher, M. (1988) Inhibitory effects of atropine and adrenergic antagonists on the changes in autonomic receptors and cyclic nucleotides of rat parotid and submandibular glands caused by sympathetic nerve stimulation. Journal of the Autonomic Nervous System, 22(1), 23–30. DOI
- Al-Manei, K., Almotairy, N., Bostanci, N., Kumar, A., Grigoriadis, A. (2020) Effect of chewing on the expression of salivary protein composition: a systematic review. Proteomics. Clinical Applications, 14(3), e1900039. DOI
- Proctor, G.B. (2016) The physiology of salivary secretion. Periodontology 2000, 70(1), 11–25. DOI
- Dolejší, E., Szánti-Pintér, E., Chetverikov, N., Nelic, D., Randáková, A., Doležal, V., Kudová, E., Jakubík, J. (2021) Neurosteroids and steroid hormones are allosteric modulators of muscarinic receptors. Neuropharmacology, 199, 108798. DOI
- Thaiss, C. A., Zmora, N., Levy, M., Elinav, E. (2016) The microbiome and innate immunity. Nature, 535(7610), 65–74. DOI
- Cua, D.J., Tato, C.M. (2010) Innate IL-17-producing cells: the sentinels of the immune system. Nature reviews. Immunology, 10(7), 479–489. DOI
- Günther, J., Seyfert, H.M. (2018) The first line of defence: insights into mechanisms and relevance of phagocytosis in epithelial cells. Seminars in Immunopathology, 40(6), 555–565. DOI
- Naafs, M. (2018) Oral mucosal immune suppression, tolerance and silencing: a mini-review. Modern Approaches in Dentistry and Oral Health Care, 1, 001–012. DOI
- Skrypnyk, M., Yatsenko, T., Riabets, O., Salama, Y., Skikevych, M., Osada, T., Tobita, M., Takahashi, S., Hattori, K., Heissig, B. (2024) Interleukin-10 induces TNF-driven apoptosis and ROS production in salivary gland cancer cells. Heliyon, 10(11), e31777. DOI
- Yu, J.C., Khodadadi, H., Baban, B. (2019) Innate immunity and oral microbiome: a personalized, predictive, and preventive approach to the management of oral diseases. The EPMA Journal, 10(1), 43–50. DOI
- Akimbekov, N.S., Digel, I., Yerezhepov, A.Y., Shardarbek, R.S., Wu, X., Zha, J. (2022) Nutritional factors influencing microbiota-mediated colonization resistance of the oral cavity: A literature review. Frontiers in Nutrition, 9, 1029324. DOI
- Skrypnyk, M., Petrushanko, T., Neporada, K., Vynnyk, N., Petrushanko, V., Skrypnyk, R. (2022) Colonization resistance of oral mucosa in individuals with diverse body mass index. Journal of Stomatology, 75, 171-175. DOI
- Shapiro, H., Thaiss, C.A., Levy, M., Elinav, E. (2014) The cross talk between microbiota and the immune system: metabolites take center stage. Current Opinion in Immunology, 30, 54–62. DOI
|